
ANATOMÍA Y FISIOLOGÍA PULMONAR (II)
Tras un pequeño periodo de espera en el que esperamos hayas tenido tiempo para leer la primera parte de esta Scubacrónica, volvemos para entregaros la segunda parte de esta serie dedicada a la fisionomía pulmonar.
Seguramente muchos de los conceptos que has leído hasta ahora son novedosos o han cambiado tu percepción sobre la anatomía pulmonar. Es normal, un curso de buceo está concebido para que todas las personas que quieren empezar a practicar esta actividad recreativa aprendan unos conceptos básicos encaminados a garantizar seguridad y confianza. Empezar una clase con temas tan extensos que pudieran parecer demasiado complicados seguramente echaría para atrás a muchos aspirantes.
Y eso que este artículo, que por su extensión hemos dividido en dos partes, está escrito de una manera muy sencilla, llana y directa, de muy fácil asimilación. Casi se puede decir que es una transcripción de una conversación entre buceadores en barra de bar.
La comprensión de la anatomía y fisionomía pulmonar nos ayudará a entender mucho mejor otros temas relacionados, como por ejemplo, el consumo de aire durante la inmersión y las formas de reducirlo, pero eso, será otra Scubacrónica. Hasta entonces, esperamos que lo que vas a leer a partir de ahora te siga resultando interesante.

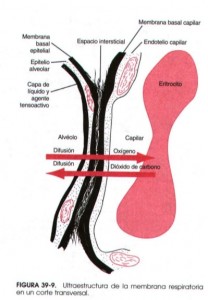
Difusión de gases por la membrana respiratoria. Las paredes alveolares son extremadamente delgadas y en su interior hay una red de pequeños vasos (capilares) interconectados como una inmensa sábana que rodea a los alveolos. El intercambio gaseoso entre el aire y la sangre se produce a través de las membranas que los separan, que en su conjunto se conocen como membrana respiratoria. En la figura inferior, vemos una imagen de microscopio electrónico de la red de capilares y la ultra-estructura de la membrana en corte transversal.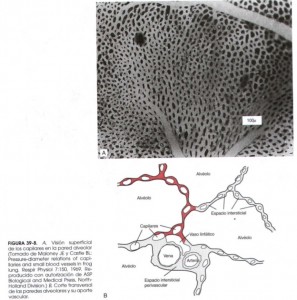
Sin entrar en la descripción de las capas que la componen, el espesor medio es de 0,6 micras (0,0006 mm). Se ha calculado que esta “alfombra” tiene una superficie de 70 metros cuadrados, lo que explica la rapidez del intercambio gaseoso. La cantidad total de sangre que hay en los capilares es de unos 100-140 ml. Imaginaros extender un vaso de agua por un salón de 70 m2; eso os dará una idea de lo fina que es la capa que se forma y lo fácil que es el intercambio. Además el diámetro de los capilares es tan pequeño que los glóbulos rojos, encargados del transporte de oxígeno, prácticamente tienen que deformarse para pasar por ellos, con lo que contactan con las paredes de los apilares haciendo más pequeña la distancia de difusión y por tanto más fácil.
Hay cuatro factores que determinan la rapidez y efectividad del paso de gases por la membrana respiratoria y cuya variación dificulta el intercambio. Vamos a verlos:
1. Espesor de la membrana: se entiende que cualquier factor que aumente el espesor de la membrana va a dificultar el intercambio. En las fibrosis pulmonares, por ejemplo, la membrana está muy engrosada y en el edema pulmonar está hinchada por líquido, que también está presente en el alveolo.
2. Área de la superficie de membrana: su reducción empeora proporcionalmente el intercambio. En el enfisema pulmonar hay muchos alveolos que han “estallado”, juntándose unos con otros. Lógicamente la superficie de una esfera grande es mucho menor que la que resulta de la suma de varias esferas pequeñas que ocupan su mismo espacio.
3. El coeficiente de difusión: Que es una constante para cada gas. Ya vimos como el CO2 difunde 20 veces mejor que el oxígeno (va a “escape libre”).
4. La diferencia de presión a través de la membrana: El famoso gradiente de presión entre las presiones parciales del gas en el alveolo y en la sangre capilar pulmonar. De los cuatro factores, éste es el más importante para nosotros, ya que es el único que puede variar de un momento para otro.
Capacidad de difusión de la membrana respiratoria. Vamos a hacer unos números para fijar este concepto. Definimos la capacidad de difusión como el volumen de gas que se difunde a través de la membrana por minuto para una diferencia de presión parcial de 1 mmHg (1 milímetro de mercurio).
En el caso del oxígeno, en condiciones de reposo la capacidad es de 21 ml/min/mm Hg. La diferencia media de presión parcial de oxígeno entre capilar y alveolo es de 11 mmHg. Multiplicando ambas cifras obtenemos 230 ml de oxígeno difundiendo hacia la sangre, que es aproximadamente la tasa de consumo de oxígeno del organismo en reposo (la “gasolina” que gastamos al ralentí).
 Durante el ejercicio intenso se aumenta el flujo sanguíneo y la ventilación alveolar, pero también aumenta la capacidad de difusión del oxígeno (hasta x3). ¿Cómo se consigue esto? Recordad el diagrama de los volúmenes respiratorios en un capítulo anterior. La respiración normal abre un pequeño número de alveolos, pero disponemos de 3 litros más que no usamos (capacidad de reserva inspiratoria). Cuando se toca el acordeón flojito se usa solo la parte media del fuelle; cuando se toca fuerte se abre y cierra casi por completo. Pues de la misma manera, nuestra “alfombra” normalmente está arrugada, ocupando solo la tercera parte del salón, pero cuando hace falta se estira. La utilización de la musculatura accesoria aumenta la presión negativa pleural y distiende más los pulmones, abriendo alveolos que permanecían cerrados (un fenómeno llamado “reclutamiento alveolar”). Me recuerda un coche que tenía mi padre con dos carburadores. El segundo funcionaba cuando pisabas fuerte el acelerador, aumentando la potencia (y el consumo, claro).
Durante el ejercicio intenso se aumenta el flujo sanguíneo y la ventilación alveolar, pero también aumenta la capacidad de difusión del oxígeno (hasta x3). ¿Cómo se consigue esto? Recordad el diagrama de los volúmenes respiratorios en un capítulo anterior. La respiración normal abre un pequeño número de alveolos, pero disponemos de 3 litros más que no usamos (capacidad de reserva inspiratoria). Cuando se toca el acordeón flojito se usa solo la parte media del fuelle; cuando se toca fuerte se abre y cierra casi por completo. Pues de la misma manera, nuestra “alfombra” normalmente está arrugada, ocupando solo la tercera parte del salón, pero cuando hace falta se estira. La utilización de la musculatura accesoria aumenta la presión negativa pleural y distiende más los pulmones, abriendo alveolos que permanecían cerrados (un fenómeno llamado “reclutamiento alveolar”). Me recuerda un coche que tenía mi padre con dos carburadores. El segundo funcionaba cuando pisabas fuerte el acelerador, aumentando la potencia (y el consumo, claro).
La Relación Ventilación/Perfusión. Hasta ahora hemos hablado de la ventilación y el intercambio como si todos los alveolos y capilares funcionaran de igual manera, pero esto no es así. Hay zonas del pulmón que tienen mayor flujo de sangre que otras. El primer factor que influye es la gravedad. La sangre es un líquido y por tanto está sometida a la ley del señor Newton. Cuando estamos de pie la mayor parte del flujo pulmonar se dirige a la parte inferior de los pulmones, dejando casi “secos” los segmentos superiores. La relación entre la ventilación y la perfusión de cada zona es un concepto clave en la fisiología pulmonar. Una zona con una relación correcta (1/1) tendrá un perfecto intercambio. Si hay ventilación pero no perfusión (1/0) el aire alveolar no puede pasar a la sangre y se añade al espacio muerto. Si hay perfusión pero no ventilación (0/1) la sangre pasa sin oxigenarse produciendo el llamado shunt o cortocircuito.
Como decíamos antes, al estar de pie la zona superior de los pulmones recibe menos sangre, con una relación ventilación/perfusión de 2,5/1; en la parte inferior esa relación baja a 0,6/1 por la mayor perfusión. Por tanto, la mayor parte de la “alfombra arrugada” está en la zona superior, y durante el ejercicio es fundamentalmente la parte superior la que se estira y recibe un gran aumento de flujo sanguíneo, mejorando la capacidad de difusión.
El organismo dispone de un mecanismo de compensación para disminuir el shunt. Cuando los alveolos no reciben oxígeno suficiente para ser funcionales segregan una sustancia que de forma local cierra los capilares pulmonares de la zona, evitando que la sangre pase por esa zona que no puede oxigenarla y haciendo de esa forma que se redirija a otras zonas mejor ventiladas. Este sistema se llama vasoconstricción pulmonar hipóxica y tiene mucha importancia, cuando se utiliza excesivamente, en la generación del edema pulmonar en situaciones de hipoxia de cualquier causa, sea por respirar con bajas presiones parciales de oxígeno (altitud, uso incorrecto de trimix hipóxico a poca profundidad) o con presiones relativas bajas (uso de un esnorkel largo, reguladores duros). Aquí entraríamos en cosas como el Edema Pulmonar por Inmersión, un concepto que está cogiendo gran auge y que se diagnostica cada vez con mayor frecuencia. Dejaremos esta enfermedad para más adelante, para no liarlo demasiado ahora.
En alguna ocasión hemos discutido la vasoconstricción pulmonar hipóxica con profundidad, principalmente en referencia a la fisiología de la altitud extrema, pero eso, aunque apasionante, es otra historia.
Con esto termina la parte de Respiración. Seguiremos hablando, si os parece, del transporte de gases en la sangre y la difusión a los tejidos, considerando las particularidades del buceo, así como de la toxicidad de los diferentes gases y sus causas.
Captación del Oxígeno por la Sangre y los Tejidos. Ya tenemos nuestra molécula de oxígeno dispuesta a entrar a la sangre. ¿Cómo lo hace y cómo se puede modificar eso?. Ya dijimos que el CO2 se difunde con gran facilidad por los líquidos y tejidos. Aun así, se combina con sustancias químicas de la sangre para aumentar unas 20 veces su capacidad de transporte. El oxígeno, sin embargo, dispone de un vehículo privilegiado: la hemoglobina presente en los eritrocitos (glóbulos rojos). Esta macromolécula tienen una enorme afinidad por el oxígeno, para el que dispone de receptores específicos, y permite que la sangre transporte entre 30 y 100 veces más cantidad de oxígeno de lo que podría hacer simplemente disolviéndolo.
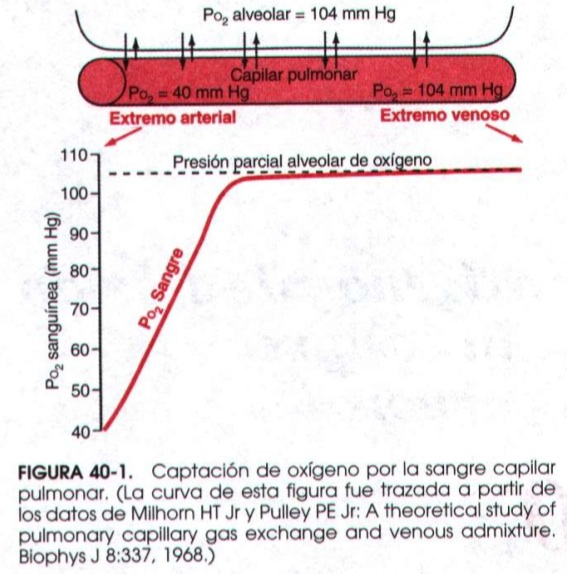 Como vimos en un capítulo anterior la presión parcial de oxígeno alveolar es de 104 mmHg. La de la sangre desaturada que proviene de los tejidos y llega a los alveolos es de uno 40 mmHg. Este gradiente hace pasar el oxígeno a los capilares, donde se une a la hemoglobina de los eritrocitos. Si veis la gráfica, las presiones se igualan cuando la sangre ha recorrido solo un tercio del capilar. El resto es tiempo de “reserva”, ahora veremos para qué.
Como vimos en un capítulo anterior la presión parcial de oxígeno alveolar es de 104 mmHg. La de la sangre desaturada que proviene de los tejidos y llega a los alveolos es de uno 40 mmHg. Este gradiente hace pasar el oxígeno a los capilares, donde se une a la hemoglobina de los eritrocitos. Si veis la gráfica, las presiones se igualan cuando la sangre ha recorrido solo un tercio del capilar. El resto es tiempo de “reserva”, ahora veremos para qué.
Durante el ejercicio intenso podemos necesitar hasta 20 veces más oxígeno que en reposo. El corazón empieza a funcionar a pleno rendimiento, aumentando tanto la frecuencia cardiaca (por tanto disminuyendo el tiempo entre latido y latido) como el volumen que expulsa en cada contracción. Como el volumen total de sangre es el mismo, el resultado lógico de lo anterior es un gran aumento de la velocidad de la sangre. Por otra parte, como explicábamos en el capítulo anterior, se abren zonas del pulmón que en reposo permanecían cerradas a la sangre para triplicar la capacidad de difusión; el resultado global es que la velocidad sanguínea aumenta entre 2 y 3 veces, acortándose en la misma medida por tanto el tiempo que tarda en pasar por el capilar pulmonar. Repasando la curva veréis que es justo el tiempo necesario para que la sangre siga consiguiendo oxigenarse completamente. Así que, aunque está disponible menos tiempo para el intercambio, se oxigena en la misma medida.
Estamos ahora en los tejidos periféricos, donde tiene lugar el proceso inverso. La presión de oxígeno en dichos tejidos es de 40 mmHg (precisamente la misma de la sangre venosa que llega hasta los pulmones procedente de los tejidos) y en el interior de las células de unos 23 mmHg. El gradiente resultante traslada el oxígeno primero de la sangre al intersticio (la zona entre células) y de él a las células. El mecanismo opuesto ocurre con el CO2 aunque, debido a su mucha mayor capacidad de difusión necesita gradientes mucho menores para difundir. Hablábamos de unos 64 mmHg de gradiente de oxígeno en los pulmones y hay unos 55 mmHg de gradiente en tejidos periféricos (esa pequeña diferencia requeriría una larga explicación que creo que no viene al caso). En el caso del CO2 el gradiente es de solo 5 mmHg entre los 40 mmHg sangre arterial y los 45 en tejido periférico. Con eso poco le basta porque difunde con gran facilidad.
 La hemoglobina también produce un sistema de compensación que hace que grandes modificaciones de la presión de oxígeno y de CO2 sean amortiguadas con pequeñas variaciones de la presión. Es algo complejo de explicar en detalle su funcionamiento (habría que hablar de la curva de disociación de la hemoglobina y sus modificaciones) y no creo que sea necesario, pero quedaros con el concepto para más adelante.
La hemoglobina también produce un sistema de compensación que hace que grandes modificaciones de la presión de oxígeno y de CO2 sean amortiguadas con pequeñas variaciones de la presión. Es algo complejo de explicar en detalle su funcionamiento (habría que hablar de la curva de disociación de la hemoglobina y sus modificaciones) y no creo que sea necesario, pero quedaros con el concepto para más adelante.
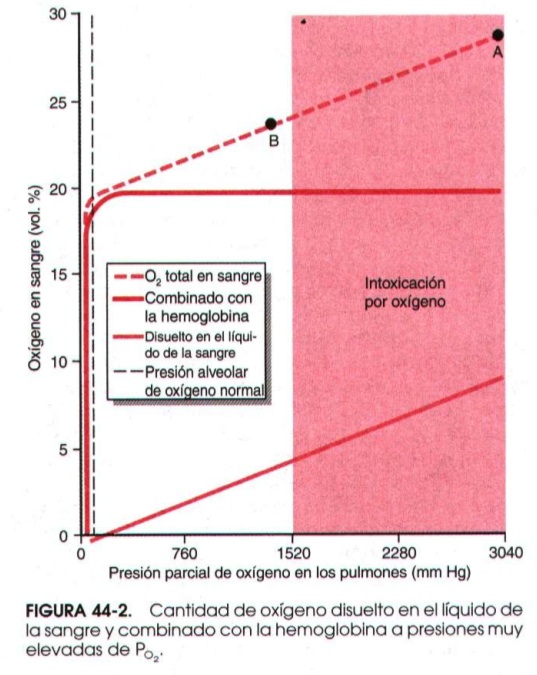
Aire enriquecido, EAN, Nitrox. Decíamos que la gran mayoría del oxígeno se transporta en la hemoglobina y solo una pequeña cantidad disuelto en sangre (en torno al 3%). La hemoglobina, respirando aire ambiente, se satura en un 95-97% de oxígeno, por lo que el hecho de respirar oxígeno a mayores concentraciones apenas puede aumentar la cantidad ligada a la hemoglobina hasta el 100%. Respirando concentraciones mayores como en el Nitrox se obtienen mayores presiones parciales de oxígeno en el alveolo, lo que genera mayor gradiente y más paso a la sangre. Como vemos, ese exceso de oxígeno casi no tiene sitio en la hemoglobina, que apenas le puede ofrecer receptores libres, así que se disuelve en la sangre hasta equilibrar la presión y por tanto pasa a los tejidos también en mayor medida al llegar a ellos. Cuando esta cantidad es excesiva y la presión tisular de oxígeno alcanza niveles muy altos se produce la famosa “intoxicación por oxígeno” con las temidas convulsiones y daño nervioso que como sabéis es el gran peligro del Nitrox mal utilizado.
Así como la enfermedad descompresiva tiene una frontera difícil de establecer con precisión (cuántas burbujas, de qué tamaño y a qué zonas van a ir son demasiadas variables), en este caso es mucho más fácil poner ese límite porque estamos hablando de un simple sistema de presiones y de reacciones químicas. Para que se desencadene la toxicidad hace falta una presión determinada, que puede ser ligeramente variable entre individuos y afectarse por condicionamientos como el ejercicio, pero dentro de unos márgenes estrechos.
Así que manteniéndonos razonablemente lejos de esa presión, que nuestras tablas y ordenadores pueden calcular con precisión, podemos estar tranquilos.
En la gráfica podéis ver como al ir aumentando la presión de oxígeno inspirada (siempre nos referiremos a este concepto, que para nosotros los buceadores es fácilmente calculable como fracción de oxígeno de la botella x presión de profundidad) la hemoglobina rápidamente se satura y lo que va aumentando es el oxígeno disuelto (la gráfica está en mmHg, recordad 1 atm=760 mmHg). Una persona que respira en reposo 4 atmósferas de oxígeno (como bajar con EAN40 a 90 metros) desarrolla convulsiones en 30-60 minutos. Pero esto disminuye con el ejercicio a valores menores por una mayor susceptibilidad de los tejidos, luego veréis por qué.
 En condiciones normales el metabolismo transforma parte del oxígeno en formas más activas, denominadas radicales libres, como el superóxido o el peróxido de hidrógeno. Esos radicales libres, de los que tanto se habla ahora en cosmética y en productos milagro, tienen un efecto muy destructivo sobre las membranas celulares si están presentes en cantidades elevadas, especialmente sobre las neuronas y el resto de tejidos nerviosos. Pero el organismo se encarga de mantenerlos dentro de niveles controlados al inactivarlos por medio de enzimas. Esta capacidad de destrucción de los radicales libres tiene un límite: por encima de 2 atmósferas de oxígeno se satura; no hay suficientes enzimas. Ese es el límite de presión de oxígeno que podemos tolerar de forma aguda sin toxicidad. Pero como es un límite aproximado, por seguridad disminuimos hasta 1,4 (1,6 en determinadas situaciones) cifra que se ha demostrado segura. No nos podemos permitir arriesgarnos a sufrir una convulsión bajo el agua bajo ningún concepto. El límite de 1,4 es seguro y debemos poner toda nuestra atención en respetarlo.
En condiciones normales el metabolismo transforma parte del oxígeno en formas más activas, denominadas radicales libres, como el superóxido o el peróxido de hidrógeno. Esos radicales libres, de los que tanto se habla ahora en cosmética y en productos milagro, tienen un efecto muy destructivo sobre las membranas celulares si están presentes en cantidades elevadas, especialmente sobre las neuronas y el resto de tejidos nerviosos. Pero el organismo se encarga de mantenerlos dentro de niveles controlados al inactivarlos por medio de enzimas. Esta capacidad de destrucción de los radicales libres tiene un límite: por encima de 2 atmósferas de oxígeno se satura; no hay suficientes enzimas. Ese es el límite de presión de oxígeno que podemos tolerar de forma aguda sin toxicidad. Pero como es un límite aproximado, por seguridad disminuimos hasta 1,4 (1,6 en determinadas situaciones) cifra que se ha demostrado segura. No nos podemos permitir arriesgarnos a sufrir una convulsión bajo el agua bajo ningún concepto. El límite de 1,4 es seguro y debemos poner toda nuestra atención en respetarlo.
Hay otra toxicidad por oxígeno que no nos afecta tanto: la pulmonar. Se puede estar respirando indefinidamente 1 atmósfera de oxígeno (por ejemplo, un paciente intubado que precisa oxígeno al 100%) sin que aparezca toxicidad nerviosa. Bueno, salvo en los casos de los recién nacidos, donde la retina es especialmente sensible. De ahí que hace años, cuando esto no se conocía, aparecieran muchos casos de ceguera infantil en prematuros tras permanecer en incubadoras con oxígeno puro (Stevie Wonder, por ejemplo).
Sin embargo, a las 12 horas a 1 atmósfera de oxígeno se comienza a desarrollar congestión de vías respiratorias, edema pulmonar y atelectasias (pérdida de aire y compactación de una zona del pulmón) por destrucción de las células de revestimiento de bronquios y alveolos, que son las que reciben de forma directa las altas cantidades de oxígeno sin que la sangre y la hemoglobina las compensen. En condiciones normales de buceo, incluso con muchas inmersiones sucesivas, no se presenta este problema ya que el tiempo entre una y otra permite recuperar la normalidad.
Otros gases. CO2 y su toxicidad. Este es un tema que siempre suscita controversia. El CO2, como hemos dicho en múltiples ocasiones, no tiene ninguna dificultad para difundir ni por los tejidos ni por la membrana pulmonar. Su producción es proporcional al metabolismo, y por tanto al nivel de ejercicio, y su eliminación es proporcional a la ventilación siempre y cuando el aire espirado salga al ambiente (o al agua).
En el caso de los rebreathers eso no es así. El aire espirado recircula y volvemos a inspirarlo, recargado con un poco de oxígeno para compensar el consumo metabólico y eliminado el CO2 por la cal sodada. Si este mecanismo falla, por ejemplo si la cal se satura completamente de CO2 y ya no puede fijar más, el nivel de CO2 inspirado irá creciendo progresivamente, disminuyendo el gradiente pulmonar y por tanto aumentando su nivel en la sangre; el círculo se cierra ya que los tejidos siguen produciéndolo y nosotros seguimos inspirándolo.
Cuando la presión de CO2 llega a 80 mmHg la situación se hace intolerable y el sistema respiratorio termina por deprimirse, complicado por la acidosis que se produce así como por la narcosis. Es el famoso tema de las apneas breves que no tiene gran impacto en los niveles de CO2 y su único peligro es realizar ascensos importantes mientras se realizan con el consiguiente riesgo de barotrauma. Otra cosa es que sean absurdas para disminuir el consumo de aire, que lo son. El consejo de no hacerlas nunca es correcto, aunque exagerado en mi opinión. Es como lo del alcohol y el embarazo: la única dosis segura es cero, lo que no quiere decir que una embarazada no se pueda tomar una caña.
N2. Nitrógeno. Hasta ahora hemos hablado mucho del oxígeno y el dióxido de carbono y nos habíamos olvidado de nuestro viejo enemigo. A nivel del mar, la cantidad de nitrógeno disuelto en el cuerpo es de casi un litro, consecuencia de que estamos respirando todo el rato un 79% de este gas. Este nitrógeno no se metaboliza. Cuando respiramos aire a presión la cantidad de nitrógeno disuelto en los líquidos corporales va aumentando y al cabo de varias horas se satura completamente igualando la presión tisular a la inspirada (fundamento de las cámaras de compresión a profundidad utilizadas para trabajos continuados a grandes profundidades). Mientras nos mantenemos en el ambiente de alta presión el nitrógeno no es un problema, pues está disuelto.
Igualmente cuando volvemos a la superficie el proceso de eliminación tarda bastante tiempo en producirse. De ese litro de nitrógeno más de la mitad está disuelto en la grasa corporal, a pesar de que dicha grasa constituye solo el 15% de un cuerpo normal. ¿Por qué? Pues debido a que el nitrógeno es 5 veces más soluble en grasa que en el agua (lipofilia). Lógicamente una persona obesa absorbe nitrógeno con mayor rapidez y en mayor cantidad por lo que es más propensa a los problemas de él derivados.
Al llegar a la saturación completa, el volumen físico que ocupa el nitrógeno en el cuerpo sigue siendo el mismo que en superficie (1 litro), pero por la conocida ley de Boyle-Mariotte de los gases, de la que ya hemos hablado (presión x volúmen = constante) corresponde a 1 x presión de profundidad (por ejemplo, 4 litros de gas comprimidos en 1 litro de volumen para 30 metros).
Decíamos que se precisan varias horas para que se equilibren las presiones de nitrógeno entre pulmones y tejidos. En la sangre ese proceso es más rápido (menos de 1 hora) pero en la grasa, que recibe cinco veces más nitrógeno y tienen menos vascularización que otros tejidos, se necesitan varias horas más.
Estos dos factores son los que, como sabéis, influyen en la carga de nitrógeno. La profundidad por su presión (y por tanto la cantidad máxima de nitrógeno a saturar) y el tiempo por el lento trasvase de nitrógeno a la grasa. No vamos a hablar ahora de compartimentos lentos y rápidos, que nos liamos.
El cómo y por qué se producen las burbujas (el cuento de la gaseosa del Open Water) y los síntomas y signos de la Enfermedad Descompresiva son de todos conocidos y no es el objeto de este texto. Quería solamente que se entendiera el concepto básico y diferencial del nitrógeno en estas circunstancias.
En cuanto al otro efecto conocido del nitrógeno, la narcosis, otro tanto. Sabéis que se produce a partir de los 30 metros (algunos hablan de 36 metros en concreto), que presenta una gran variabilidad individual, que su peligro es el mal juicio que puede producir la euforia y que se corrige inmediatamente al ascender. Es un efecto puramente narcótico del sistema nervioso central y no tiene mayor transcendencia fisiológica. Ciertamente hay personas más sensibles que otras, pero a todos nos afecta en mayor o menor medida. Es un problema de graduación. Yo nunca he sido consciente de ella, a pesar de haber bajado muchas veces de los 35 metros, pero estoy seguro de que si realizo un test suficientemente bueno para evaluar mis capacidades intelectuales superiores a esa profundidad mis resultados serían peores que en superficie. Y el vuestro también.
H2. Helio. Como todos los que usáis Trimix sabéis, la utilización de Helio en la mezcla de gases de la botella cumple un objetivo múltiple para el buceo a gran profundidad: disminuye tanto la concentración de nitrógeno como la de oxígeno (en el trimix hipóxico) para tratar de evitar los efectos adversos y las toxicidades de ambos gases a presión elevada:
– disminuir el efecto narcótico del nitrógeno (el del helio es la quinta parte)
– disminuir su disolución tisular (la mitad) y mejorar la eliminación
– disminuir la resistencia de las vías aéreas a la respiración, ya que el nitrógeno muy comprimido es bastante denso y aumenta el trabajo respiratorio, mientras el helio es siete veces menos denso (el peso molecular del N2 es 28, el del He2 es 4).
– disminuir la concentración de oxígeno para que, al multiplicarla por una presión de profundidad elevada, nos mantenga una presión alveolar de oxígeno por debajo de la tóxica.
Control de la Respiración
Como sabemos, la respiración normal es un acto completamente inconsciente. No tenemos que pensar ni realizar una acción voluntaria para respirar, no decidimos cuándo ni cuánto respiramos. Nuestro “sistema operativo” se encarga de ello. Para hacerlo dispone de un chip, el Centro Respiratorio, y de unos sensores de varios tipos que le suministran la información necesaria para la toma de decisiones.
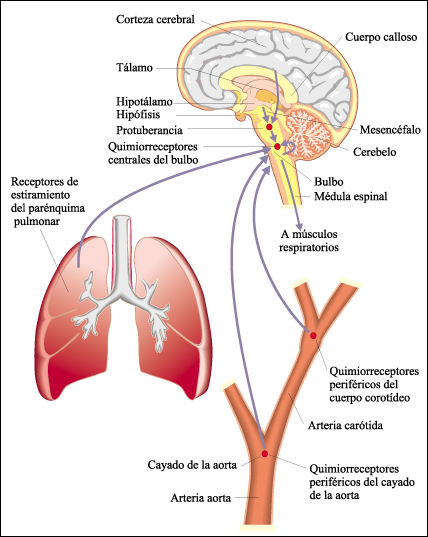 El Centro Respiratorio está compuesto por varios grupos de neuronas localizadas en el bulbo raquídeo y la protuberancia, que corresponde a la zona baja del cerebro, cerca de la conexión de éste con la médula espinal. No vamos a entrar en la descripción detallada de los grupos neuronales y sus funciones.
El Centro Respiratorio está compuesto por varios grupos de neuronas localizadas en el bulbo raquídeo y la protuberancia, que corresponde a la zona baja del cerebro, cerca de la conexión de éste con la médula espinal. No vamos a entrar en la descripción detallada de los grupos neuronales y sus funciones.
Los receptores son de tres tipos:
1. Quimiorreceptores periféricos: fundamentalmente en la aorta y las carótidas. Informan de parámetros químicos de la sangre al centro respiratorio.
2. Barorreceptores que informan de la presión.
3. Receptores pulmonares que informan sobre el grado de distensión pulmonar, localizados en las porciones musculares de los bronquios.
La señal que pone en marcha la inspiración se origina por tanto en un grupo de esas neuronas y tiene una forma de “rampa”. En la respiración normal, la inspiración comienza débilmente y crece durante unos 2 segundos. Cesa de forma repentina los 3 segundos siguientes, lo que interrumpe la estimulación del diafragma y permite la retracción elástica de la pared torácica y los pulmones, o sea la espiración. Luego el ciclo vuelve a comenzar. Ese proceso de 5 segundos nos da la frecuencia respiratoria normal (12 ciclos por minuto). Pensad nuevamente en un acordeón, esta vez con un lateral apoyado en el suelo y nuestra mano en el lado contrario. El proceso activo es tirar de nuestra mano hacia arriba para hincharlo. Luego se deja bajar la mano y la gravedad hace el trabajo. Pues aquí, puesto del revés, es lo mismo, salvo que en vez de la gravedad son las fuerzas elásticas las que provocan la espiración. Esto lo comentamos en los capítulos iniciales.
Hay dos formas de controlar esa rampa inspiratoria: aumentando la velocidad del ascenso (llenando los pulmones de manera más rápida) o aumentando el punto límite, en el que cesa la rampa. Esta última es la forma habitual de control de la frecuencia respiratoria: cuanto antes cese la rampa, más corta será la inspiración y mayor la frecuencia (también se acorta la espiración). Este control lo realiza una zona concreta denominada “centro neumotáxico”, cuya función puede modificar la frecuencia respiratoria por el mecanismo descrito desde 3-5 hasta 30-40 respiraciones por minuto.
Hay otro grupo de neuronas denominadas “grupo respiratorio ventral” que serían como el “turbo”. No intervienen en el control de la respiración normal, pero cuando se precisa mucho mayor trabajo se ponen en marcha, estimulando especialmente los músculos abdominales, que recordaréis que dijimos que son los principales responsables de la respiración forzada. Los receptores pulmonares de los que hablábamos al principio transmiten señales de distensión pulmonar que interrumpen la rampa inspiratoria. Este fenómeno se llama “reflejo de insuflación de Hering-Breuer”, que en los humanos se pone en marcha cuando el volumen corriente se triplica (1,5 litros). Si probáis a hacer inspiraciones lentas pero muy profundas notaréis una especie de “fuerza interior” que intenta interrumpirlas y hay que seguir “tirando” voluntariamente para vencerla. Los apneistas saben mucho de esto.
El asunto ahora es: ¿cómo se produce el control de la respiración cuando cambian las condiciones? Acabamos de ver el reflejo de insuflación, pero el meollo está en el control químico.
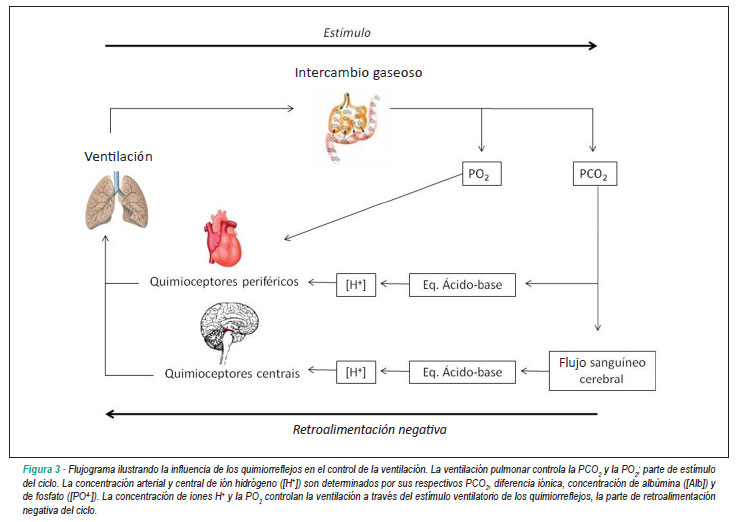 El nivel de oxígeno y el de CO2 son los principales factores que intervienen, pero no los únicos. El oxígeno no tiene un efecto directo importante en el centro respiratorio, sino que actúa a través de los quimiorreceptores que citamos anteriormente, los cuales transmiten la señal al cerebro. El CO2, por el contrario, es un potente estimulador directo del centro respiratorio, cuyo efecto es mayor que el del oxígeno. Esta estimulación se produce en una pequeña área “quimiosensible” que activa los grupos de neuronas descritos anteriormente. La descripción detallada de los procesos bioquímicos por los que el CO2 estimula estas neuronas, aunque interesante, sería demasiado extensa. Quedaros con la idea de que el CO2 elevado es el estímulo más potente del centro respiratorio, concepto al que volveremos en varias ocasiones.
El nivel de oxígeno y el de CO2 son los principales factores que intervienen, pero no los únicos. El oxígeno no tiene un efecto directo importante en el centro respiratorio, sino que actúa a través de los quimiorreceptores que citamos anteriormente, los cuales transmiten la señal al cerebro. El CO2, por el contrario, es un potente estimulador directo del centro respiratorio, cuyo efecto es mayor que el del oxígeno. Esta estimulación se produce en una pequeña área “quimiosensible” que activa los grupos de neuronas descritos anteriormente. La descripción detallada de los procesos bioquímicos por los que el CO2 estimula estas neuronas, aunque interesante, sería demasiado extensa. Quedaros con la idea de que el CO2 elevado es el estímulo más potente del centro respiratorio, concepto al que volveremos en varias ocasiones.
Este estímulo se va agotando con el tiempo. Al cabo de unas cuantas horas disminuye su efecto. Las personas con enfermedad pulmonar crónica que ventilan poco retienen CO2 de forma crónica y en ellas este efecto está reducido a una quinta parte de su fuerza, tomando el control de la respiración el siguiente elemento en la lista: el oxígeno. Por eso a estas personas, a pesar de que tienen hipoxemia crónica (bajos niveles de oxígeno en sangre) no se les puede administrar oxígeno a altas concentraciones: su estímulo número uno, el CO2 alto, está anulado. Si aumentamos demasiado sus bajos niveles de oxígeno (estímulo número dos para nosotros, número uno para ellos), respiran menos y retienen aún más CO2. Es un efecto aparentemente paradójico (darles mucho oxígeno les lleva a empeorar la respiración) pero tiene una clara base fisiológica.
¿Por qué es mucho mayor el control sobre el CO2 que sobre el oxígeno, cuando es éste último el combustible básico de nuestro metabolismo? Si recordáis lo dicho en capítulos anteriores, la función de amortiguación de la hemoglobina hace que en casi cualquier situación los niveles de oxígeno estén asegurados. Por tanto, excepto en condiciones especiales, el aporte de oxígeno es adecuado a pesar de grandes variaciones de la ventilación pulmonar (entre menos de la mitad y 20 veces más de lo normal). Sin embargo la presión de CO2 es un indicador exacto de la ventilación. La evolución ha hecho que el parámetro más fiable de lo que el centro respiratorio quiere medir sea el principal responsable de su control. El oxígeno entra en la foto solamente cuando su presión disminuye por debajo de 60-70 mmHg, o sea cuando nos metemos en el rango de la Insuficiencia Respiratoria
El artículo “Anatomía, Fisiología Pulmonar y Buceo” que reproducimos en esta serie de tres “scubartículos” fueron publicados el 9 de marzo de 2010 en Forobuceo. Contamos con la autorización expresa de su autor “Decoman”, para reproducirlos. Agradecemos, por tanto, al autor, su amabilidad al permitir la difusión de estos artículos y reconocemos el esfuerzo y la calidad atesorada en la realización de este trabajo.
Zona de inMersión: La seguridad nos permite detectar el error. La reflexión indica qué hay que hacer. La destreza práctica indica cómo hay que hacerlo. La formación y la experiencia son necesarias ganar seguridad.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}